

Długość: do kilkunastu centymetrów, SVL (dł. bez ogona) niecałe 7 cm
Masa: kilkanaście g
Czas: santon?-kampan, ok. 85-70 mln lat temu
Miejsce: centralna Azja, Mongolia i Kazachstan
Klasyfikacja:
Diapsida
Lepidosauromorpha
Lepidosauria
Squamata
Lacertiformes?
Slavoia
S. darevskii
Wstęp:
Skamieniałe kości Slavoia darevskii zostały odkryte w czasie polsko-mongolskich wypraw paleontologicznych w latach sześćdziesiątych i siedemdziesiątych ubiegłego wieku. Zgromadzono wtedy kolekcję składającą się z kilkudziesięciu okazów tej jaszczurki, były to głównie czaszki. Opisał je Andrzej Sulimski na łamach Palaeontologia Polonica w 1984 roku. W pracy tej znalazł się dokładny opis anatomiczny ze szczególnym naciskiem na budowę czaszki. Nadana przez niego nazwa rodzajowa wywodzi się od drugiego imienia młodszego brata autora, gatunkowa od nazwiska profesora – prof. Ilji S. Darewskiego z Sankt Petersburga.
Występowanie:
W trakcie polsko-mongolskich wypraw na pustynię Gobi (1964-1971) znaleziono kilkadziesiąt okazów tej jaszczurki. Sulimski (1984) doniósł, że pochodziły one z trzech stanowisk: Chulsan, Nemegt (formacja Barun Gojot) i Chermin Caw II (czerwone warstwy Chermin Caw). W przypadku Nemegt autor miał prawdopodobnie na myśli niższe warstwy Nemegt (Lower Nemegt Beds), które są obecnie nazywane formacją Barun Gojot (Gradziński i Jerzykiewicz, 1974). Wśród znalezisk z wyższych warstw Nemegt (Upper Nemegt Beds) nie są wymieniane żadne jaszczurki (Kielan-Jaworowska i Dovchin, 1969; Kielan-Jaworowska i Barsbold, 1972; Gradziński, 1970). Obecnie Chermin Caw również jest zaliczane do formacji Barun Gojot (Gao i Norell, 2000). Zatem wszystkie okazy zebrane przez polskich paleontologów w czasie tamtych wypraw pochodziły z jednej formacji.
Od lat siedemdziesiątych również inne ekspedycje znajdowały jaszczurki należące do tego gatunku. Według The Paleobiology Database (2011) skamieniałości S. darevskii z pustyni Gobi w Mongolii są w pięciu różnych kolekcjach na świecie, przy czym jest to zapewne liczba niepełna. Wiele okazów znaleźli badacze amerykańscy. Prowadzili oni intensywne poszukiwania nie tylko na terenie Chulsan i Chermin Caw, ale także w formacji Dżadochta. Ze wszystkich tych miejsc udało im się zebrać sporą liczbę kredowych jaszczurek. Jednak S. darevskii nie została znaleziona na terenie formacji Dżadochta, z wyjątkiem stanowiska Uchaa Tołgod (Ryc. 1) (Gao i Norell, 2000). Kordikova i in. (2001) zidentyfikowali kość zębową należącej do S. cf. darevskii z formacji Bostobe w Kazachstanie (stanowisko Szach-Szach).
Podsumowując, gatunek ten występował prawdopodobnie na terenie całej centralnej Azji – został znaleziony na terenie formacji Barun Gojot (kilka miejsc w Chulsan, Chermin Caw I i II, niższe warstwy Nemegt), formacji Dżadochta (tylko Uchaa Tołgod) w Mongolii (Sulimski, 1984; Gao i Norell, 2000) i formacji Bostobe w Kazachstanie (Kordikova i in., 2001). Jak na gatunek kopalnego kręgowca, jest dosyć liczny i szeroko rozpowszechniony.
Wiek stanowisk:
Formacja Barun Gojot i Dżadochta datowane są na kampan (około 83,5 do 70,6 miliona lat temu). Datowanie stanowisk mongolskich nie jest jednak zbyt dokładne ze względu na brak możliwości datowania radiometrycznego i bezpośredniej korelacji z osadami morskimi (Gao i Norell, 2000). Dlatego wiek formacji mongolskich oszacowano głównie na podstawie podobieństwa fauny kręgowców do innych stanowisk na świecie, co nie jest precyzyjną metodą (Gradziński i Jerzykiewicz, 1974). Formacja Dżadochta może być nieco starsza lub równowiekowa Barun Gojot (Gao i Norell, 2000). Na Barun Gojot zalegają zgodnie górne warstwy Nemegt, które mogą być już mastrychckiego wieku (Gradziński i Jerzykiewicz, 1974). Formacja Bostobe jest datowana na santon lub wczesny kampan i pochodzący stamtąd okaz może być najstarszym przedstawicielem rodzaju (Kordikova i in., 2001; Nessov i in., 1998).
Geologia stanowisk:
Barun Gojot składa się z klastycznych osadów kontynentalnych. Dolna część z warstwowaniem skośnym jest interpretowana jako osady wydmowe, poprzeplatane osadami wodnymi, które tworzyły się w strefach między-wydmowych. W formacji dominują miąższe piaskowce o różnym pochodzeniu. W górnej części profilu są to piaskowce o poziomym warstwowaniu, które mogą być zapisem okresowych powodzi. Większość znalezisk jaszczurek pochodzi z luźnych brył, znajdowanych ex situ. Te, które udało się znaleźć w profilu, występowały w różnie warstwowanych piaskowcach z małymi otoczakami. Część skamieniałości pochodzi z masywnych piaskowców. Jaszczurki występują zatem w niemal całym profilu; podobnie ssaki. Natomiast skamieniałości dinozaurów, żółwi i krokodyli znajdowane były głównie w jego dolnej części (Gradziński i Jerzykiewicz, 1974).
Niższa część formacji Dżadochta składa się z osadów rzecznych, w przeciwieństwie do pozostałej części, która jest eoliczna. Dlatego Hasegawa i in. (2009) zaproponowali, by z niższej części wydzielić nową formację o nazwie Alagteeg. Jej osady charakteryzują się poziomo warstwowanymi piaskowcami i mułowcami, będącymi świadectwem piaszczystych rzek, równi zalewowych i efemerycznych zbiorników. Nad nimi zalegają gruboławicowe, skośnie warstwowane piaskowce, caliche i soczewki cienkich mułowców. Osady wskazują na środowisko pustynne z wydmami. Jeszcze wyżej w profilu (górna część formacji Dżadochta) występują bezstrukturalne piaskowce, caliche in situ, piaskowcowe egzotyczne otoczaki i duże nory. Świadczy to środowisku przypominającym step z bogatą w wapień glebą tworzącą się w półsuchym klimacie (Hasegawa i in. 2009).
W Uchaa Tołgod skamieniałości występują w niewarstwowanych piaskowcach, natomiast brak ich w piaskowcach wydmowych (Loope i in., 1998). Osady zawierające kości interpretuje się jako spływ materiału z wydm piaskowych pod wpływem ulewnych deszczy. Osad pokrył martwe lub umierające zwierzęta a niektóre pogrzebał w ich norach. W ten sposób tłumaczy się występowanie licznych szkieletów zwierząt na tym terenie (Loope i in., 1998; Gao i Norell, 2000). Zanik części warstwowania jest tłumaczona jako skutek bioturbacji – zadeptywania przez dinozaury i drążenie norek przez bezkręgowce (Loope i in., 1998). Występują także osady iluwialne i wapienie pedogeniczne. Ogólnie osady z tego stanowiska reprezentują środowisko bardziej wilgotne niż w pozostałej części formacji (Gao i Norell, 2000).
Formacja Bostobe składa się z szarych iłów i czerwonawych piasków, miejscowo występują skośnie warstwowane piaskowce. Większość skamieniałości występuje w ciemnej, bogatej w materię organiczną warstewce, 9 metrów od spągu profilu. Fauna jest reprezentowana przez skorupiaki, rekiny, ryby, płazy, jaszczurki, żółwie i dinozaury. Szatę roślinną tworzyły głównie sekwoje, ale także cypryśniki, wiązy, dęby, wierzby i topole. Osady formacji interpretuje się jako estuariowe (Kordikova i in., 2001). Znalezisko pojedynczej kości Slavoia jest wyjątkowe i nie jest wystarczającym dowodem o związku tego zwierzęcia z takim środowiskiem.
Środowisko życia Slavoia darevskii:
Formacja Barun Gojot i stanowisko Uchaa Tołgod były prawdopodobnie bardziej wilgotne niż pustynna Dżadochta. Więcej w nich osadów wodnych i śladów pokrywy roślinnej (Gao i Norell, 2000). To może być środowisko występowania S. darevskii. Gao i Norell (2000) donoszą, że S. darevskii była znajdowana w luźnym piasku, podobnie jak Phrynosomimus asper. Oba gatunki musiały zajmować podobne siedliska i prawdopodobnie żyły w norach (Sulimski, 1984; Gao i Norell, 2000). W Uchaa Tołgod niektóre szkielety Parmeosaurus scutatus były znajdowane w skulonej pozycji z głową podciągniętą pod ogon. Okazy te występowały w mocniej zlityfikowanych fragmentach piaskowca. Interpretuje się takie okazy jako jaszczurki pogrzebane przyżyciowo w norach, na co wskazuje także morfologia czaszki tego gatunku, typowa dla jaszczurek kopiących w ziemi (Gao i Norell, 2000).
W przypadku S. darevskii brak na to bezpośrednich dowodów. Jednak w zbiorach Instytutu Paleobiologii PAN znajdują się szkielety w skulonej pozycji (MgR-I/8, MgR-I/99). Niewypreparowany piaskowiec, na którym spoczywają te okazy jest mocniej zlityfikowany. Gradziński i Jerzykiewicz (1974) napisali, że większość jaszczurek w Barun Gojot znaleziono w okruchach piaskowca u podnóża ścian. Nieliczne znajdowane in situ zawsze występowały w różnie warstwowanych piaskowcach, osadzone raczej w warunkach wodnych (Gradziński i Jerzykiewicz, 1974). Można przypuszczać, że znajdowane u podnóża ścian fragmenty piaskowca były bardziej zlityfikowane niż otaczający je osad i zostały z tych ścian naturalnie wypreparowane. Niektóre okazy S. darevskii mogły więc być pogrzebane w norze (Tałanda, 2011).
Budowa Slavoia darevskii:
Czaszka S. darevskii widziana z boku i góry w swym zarysie ma kształt łzy. Zwęża się ku przodowi, tył natomiast jest szeroki. Przód czaszki tego gatunku ma zwartą budowę - charakteryzuje się dość krótkimi i szerokimi kośćmi czołowymi, które łączą się ze szczękowymi. Oddzielają przez to kości nosowe od przedczołowych. Otwór ciemieniowy jest mały lub go nie ma. Uwagę zwracają zredukowane oczodoły o łezkowatym kształcie. Kości przedczołowe łączą się z zaczołowymi, które nie są zrośnięte z zaoczodołowymi, jak u niektórych jaszczurek. Kość jarzmowa ma patykowaty kształt i sięga kości łuskowej, a jej wyrostek tylny jest bardzo mały lub nie występuje. Kość łuskowa również jest dosyć długa i prosta, ale ze skrzywionym tylnym końcem bardziej przypomina kij do hokeja. Brak u niej wyrostka grzbietowego. Dół nadskroniowy jest mały, ale większy od oczodołu, zwrócony nieco ku tyłowi. Uzębienie tej jaszczurki jest dość nietypowe. Zęby mają jednolitą budowę, są cylindryczne, ostre, bez spłaszczenia i urzeźbienia, dość luźno upakowane. Były umieszczone w szczękach pleurodontycznie i wymieniały się sukcesywnie. Na kości przedszczękowej było ich 5, szczękowej 7-8, a na żuchwie 9-10. S. darevskii ma małą kość łzową, niezrośniętą z przedczołową, która mieści się w całości w oczodole. Lemiesze są bezzębne. Kości podniebienne zawijają się i łączą się szwem z lemieszami. Okno podoczodołowe jest bardzo małe. Żuchwa jest wysoka, dosyć masywna. Naprzeciw kości płatowej szeroko otwiera się rowek Meckela, jak zauważył Sulimski (1984). Autor ten nie przedstawił w swej pracy budowy mózgoczaszki, argumentując, że będzie ona przedmiotem kolejnej pracy. Nie ukazała się jednak na ten temat żadna publikacja.
Kręgosłup składa się z procelicznych (przodowklęsłych) kręgów. W odcinku szyjnym jest ich 7, a w całej przedkrzyżowej części 26. Międzyobojczyk ma kształt krzyża, z szerokimi wyrostkami bocznymi i silnym grzebieniem poprzecznym. Obojczyki są cienkie, długie, poszerzają i rozwidlają się dośrodkowo. Kość krucza jest dosyć rozszerzona. Sulimski (1984) przedstawił budowę kończyn i miednicy pobieżnie ze względu na stan zachowania i niedokończoną preparację okazów. Stwierdził, że kończyny były raczej kompletne, ale niewyspecjalizowane i o zredukowanej długości.
Paleobiologia:
Budowa obręczy barkowej sugeruje obecność silnych mięśni szyi i dobrze rozwiniętych mięśni kończyn przednich. Prawdopodobnie Slavoia była przystosowana do podziemnego trybu życia. Tę hipotezę wspiera redukcja oczodołów i silnie skostniały, masywny przód czaszki. Jaszczurka ta miała raczej słaby wzrok.
Pozycja filogenetyczna i klasyfikacja:
Sulimski (1984) zaklasyfikował Slavoia darevskii do scynkokształtnych (Scincomorpha) na podstawie następujących cech: parzyste kości nosowe i czołowe, pojedyncza kość ciemieniowa ze zredukowanym otworem, zawinięte kości podniebienne i wtórne podniebienie, osobna kość zaczołowa i zaoczodołowa, budowa kości łopatkowo-kruczej, wydłużona mózgoczaszka, struktura szczęki dolnej i generalna tendencja do redukcji kończyn. Takson ten wykazywał według niego również pewne podobieństwa do form bliżej spokrewnionych z gekonami, ale ze względu na inne cechy, uznał je za konwergentne. Sulimski widział też pewne podobieństwa S. darevskii do jaszczurek makrocefalozaurydowych i poliglifanodontowych z tych samych stanowisk, a także do podrodzin Teiinae i Gymnophtalminae. Z tą ostatnią nie wykluczał bliższego pokrewieństwa. Wątków tych jednak nie rozwinął. Ostatecznie pozostał przy stanowisku o przynależności tej jaszczurki do scynkokształtnych. Nie próbował przy tym oznaczyć jej do którejś z rodzin scynkokształtnych ze względu na mozaikę cech różnych przedstawicieli tej grupy (Sulimski, 1984).
Borsuk-Białynicka (1988) zasugerowała, że Slavoia i Globaura są prymitywnymi, pośrednimi przedstawicielami Lacertiformes i reprezentują stan cech prymitywnych dla scynkokształtnych. Gao i Fox (1991) potwierdzili to stanowisko, dodając, że Slavoia nie ma żadnych synapomorfii z Gymnophthalmidae. Borsuk-Białynicka (1991) wspomniała enigmatycznie, że Eoxanta i, być może, S. darevskii są siostrzane dla Xantusiidae i Scincidae. Alifanov zaproponował (1993), by te dwa rodzaje zaliczyć do nowej rodziny Slavoiidae i że mogą być pokrewne Eichstaettisaurus z jury Niemiec, a także do Acontiidae – podrodziny Scincidae. Niestety nie podał żadnego uzasadnienia tej opinii. Gao i Norell (2000) podali klasyfikację Alifanova (1993) w wątpliwość. Piszą, że u S. darevskii wprawdzie ectopterygoid kontaktuje się z kością podniebienną przednio i tylnie jak u Acontiidae, ale brak jest innych cech typowych dla tej grupy (np. utrata kończyn i łuku nadskroniowego, redukcja kości przedczołowej i łuskowej, zamknięty kanał Meckelego z fuzją). Gao i Norell przyznają, że zwinięcie kości podniebiennej sugeruje jakieś pokrewieństwo ze Scincidae, ale brak takich cech jak kontakt kości łuskowej i jarzmowej nie pozwalają zaliczyć tego gatunku do tej grupy. Obaj autorzy pozostawiają sprawę dokładniejszej klasyfikacji Slavoia wewnątrz scynkokształtnych za otwartą. W artykule Kordikovej i in. (2001) Slavoia za Alifanovem (1993) jest zaklasyfikowana do rodziny Slavoiidae.
Dobrze poznana osteologia i kredowy wiek czynią ze S. darevskii dogodny i ważny takson do badań filogenetycznych gadów łuskonośnych, w szczególności tych wczesnych. W analizach filogenetycznych Kearney (2003a i 2003b), których głównym celem było zbadanie pokrewieństwa Sineoamphisbaena hexatabularis i amfisben, znalazła się również S. darevskii. W tych analizach Slavoia jest blisko Adamisaurus i kladu zawierającego Sineoamphisbaena, Macrocephalosauridae i Polyglyphanodontidae. Kladogram otrzymany przez Kearney (2003a) sugeruje bliższe pokrewieństwo do Lacertidae i Teiidae niż do Scincidae i Cordylidae, podobnie jak badania Borsuk-Białynickiej (1988), jednak wyklucza ze scynkokształtnych główne beznogie grupy – węże, amfisbeny i dibamidy.
Jedną z większych analiz kladystycznych z użyciem cech morfologicznych gadów łuskonośnych była praca Conrada (2008). W tej publikacji Slavoia w obrębie scynkokształtnych znalazła się jako grupa zewnętrzna Scincophidia (nowo nazwanego kladu, zawierającego węże, amfisbeny, dibamidy i scynki). Analiza Conrada, w przeciwieństwie do poprzednich badań (Borsuk-Białynicka, 1988; Kearney, 2003a) sugeruje bliższe pokrewieństwo Slavoia do scynków i amfisben, niż do Lacertiformes. Należy jednak pamiętać, że wśród filogenetyków toczy się dyskusja, czy w ogóle scynkokształtne są monofiletyczne (Conrad, 2008 i cytowane tam prace). Dlatego można uznać pozycję filogenetyczną Slavoia darevskii za sprawę dość otwartą.
Literatura:
1. Alifanov, V. 1993. Some peculiarities of the Cretaceous and Palaeogene lizard faunas of the Mongolian People's Republic. Kaupia 3, 9-13.
2. Borsuk-Białynicka, M. 1988. Globaura venusta gen. et sp. n. and Eoxanta lacertifrons gen. et sp. n. - non-teiid lacertoids from the Late Cretaceous of Mongolia. Acta Palaeontologica Polonica 33, 211-248.
3. Borsuk-Białynicka, M. 1991. Questions and controversies about saurian phylogeny, Mongolian perspective. W: Kielan-Jaworowska, Z., Heintz N. i H. A. Nakrem, (red.). Fifth Symposium on Mesozoic Terrestrial Ecosystems and Biota, Oslo, 9-10.
4. Conrad, J. L. 2008. Phylogeny and systematics of Squamata (Reptilia) based on morphology. Bulletin of the American Museum of Natural History 310, 1-182.
5. Estes, R., de Queiroz, K. i J., Gauthier. 1988. Phylogenetic Relationships within Squamata. W: Estes, R. i G., Pregill (red.). Phylogenetic Relationships of the Lizard Families: Essays Commemorating Charles L. Camp. Stanford University Press, 119-281.
6. Gao, K.-Q. i R. C., Fox. 1991. New teiid lizards from the Upper Cretaceous Oldman Formation (Judithian) of southwestern Alberta, Canada, with a review of the Cretaceous of the Cretaceous record of teiids. Annals of the Carnegie Museum 60, 145-162.
7. Gao, K.-Q. i M. A., Norell. 2000. Taxonomic composition and systematics of Late Cretaceous lizard assemblages from Ukhaa Tolgod and adjacent localities, Mongolian Gobi Desert. Bulletin of the American Museum of Natural History 249, 1-118.
8. Gradziński, R. 1970. Sedimentation of dinosaur-bearing Upper Cretaceous deposits of the Nemegt Basin, Gobi Desert, Mongolia. Palaeontologia Polonica 21, 147-229.
9. Gradziński, R. i T., Jerzykiewicz. 1974. Sedimentation of the Barun Goyot Formation. Palaeontologica Polonica 30, 11-146.
10. Hasegawa H., Tada, R., Ichinnorov, N. i C., Minjin. 2009. Lithostratigraphy and depositional environments of the Upper Cretaceous Djadokhta Formation, Ulan Nuur Basin, southern Mongolia, and its paleoclimatic implication. Journal of Asian Earth Sciences 35, 13-26.
11. Kearney, M. 2003a. The phylogenetic position of Sineoamphisbaena hexatabularis reexamined. Journal of Vertebrate Paleontology 23, 394-403.
12. Kearney, M. 2003b. Systematics and Evolution of the Amphisbaenia (Lepidosauria: Squamata): A Phylogenetic Hypothesis Based on Morphological Evidence from Fossil and Recent forms. Herpetological Monographs 17, 1-74.
13. Kielan-Jaworowska, Z. i N., Dovchin. 1969. Narrative of the Polish-Mongolian Palaeontological Expeditions. Palaeontologia Polonica 19, 7-30.
14. Kielan-Jaworowska, Z. i R., Barsbold. 1972. Narrative of the Polish-Mongolian Palaeontological Expeditions, 1967-1972. Palaeontologia Polonica 27, 1-12.
15. Kordikova, E. G., Polly, P. D., Alifanov, V. A., Roček, Z., Gunnell, G. F. i A. O., Averianov. 2001. Small vertebrates from the Late Cretaceous and Early Tertiary of the Northeastern Aral Sea Region, Kazakhstan. Journal of Paleontology 75, 390-400.
16. Loope, D. B., Dingus, L., Swisher III, C. C. i C., Minjin. 1998. Life and death in a Late Cretaceous dune field, Nemegt basin, Mongolia. Geology 26, 27-30.
17. Nessov, L. A., Archibald, J. D., i Z., Kielan-Jaworowska. 1998. Ungulate-like mammals from the Late Cretaceous of Uzbekistan and a phylogenetic analysis of Ungulatomorpha. Bulletin of the Carnegie Museum of Natural History 34, 40-88.
18. Sulimski, A. 1984. A new Cretaceous scincomorph lizard from Mongolia. Palaeontologica Polonica 46, 143–155.
19. Tałanda, M. 2011. Biologia późnokredowej jaszczurki Slavoia darevskii na podstawie morfologii szkieletu. Niepublikowana praca magisterska. Archiwum Prac Dyplomowych Uniwersytetu Warszawskiego, 1-92.
20. Townsend, T. M., Larson, A., Louis, E. i J. R. Macey. 2004. Molecular phylogenetics of Squamata: the position of snakes, amphisbaenians, and dibamids, and the root of the squamate tree. Systematic Biology 53, 735–757.
21. Vidal, N. i S. B., Hedges. 2005. The phylogeny of squamate reptiles (lizards, snakes, and amphisbaenians) inferred from nine nuclear protein-coding genes. Comptes Rendus Biologies 328, 1000-1008.
22. Vidal, N. i S. B., Hedges. 2009. The molecular evolutionary tree of lizards, snakes, and amphisbaenians. Comptes Rendus Biologies 332, 129-139.
Ilustracje:

Zwinięty przyżyciowo młody osobnik MgR-I/112. Być może zasypany żywcem w norze.

Okaz holotypowy MgR-I/8.

Fragment szkieletu osobnika MgR-III/79. Widoczna jest dobrze rozwinięta kończyna przednia.
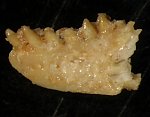
Kość zębowa z charakterystycznymi, kołkowatymi zębami.





